Opções de inscrição
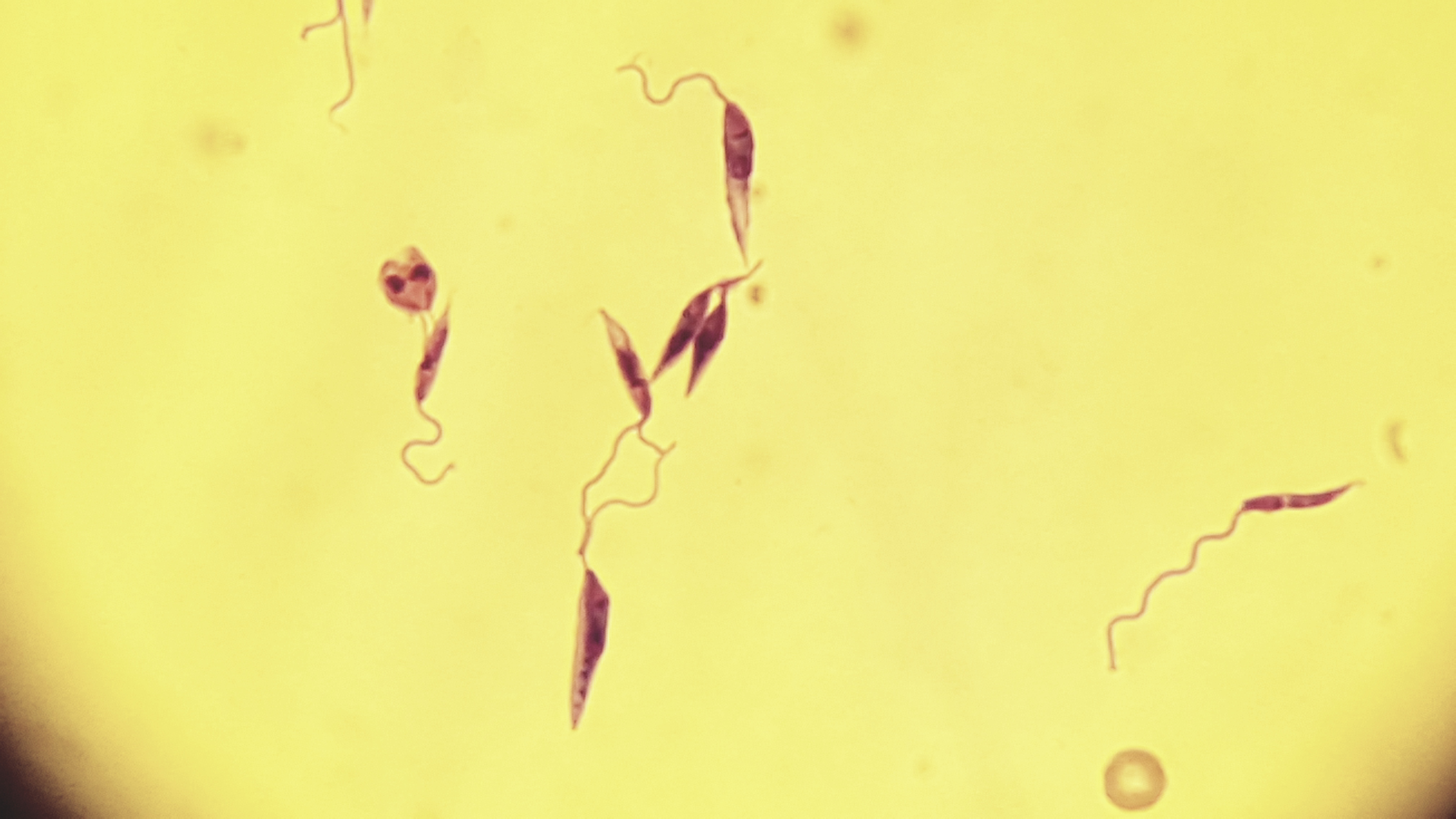
Apresentar e discutir problemas e pesquisas envolvendo os mecanismos de resposta imune, patogênese, diagnóstico, epidemiologia, biologia, tratamento e controle das leishmanioses em animais domésticos, para que o estudante desenvolva um pensamento crítico sobre o tema.
Visitantes não podem acessar este curso. Por favor faça login.