[Apostila online] Tópico 7: Organizando o armário!
| Site: | Moodle USP: e-Disciplinas |
| Curso: | 05 - Zoologia |
| Livro: | [Apostila online] Tópico 7: Organizando o armário! |
| Impresso por: | Usuário visitante |
| Data: | quinta-feira, 11 jul. 2024, 19:38 |
1. Início de Conversa

- A Escola Essencialista
Para começar a ser considerada uma prática científica, a Sistemática (ciência responsável pela classificação dos seres vivos) teve de esperar até 350 a.C., quando Aristóteles resolveu classificar a vida. Ele seguia o essencialismo platônico, no qual o que realmente importa são os traços típicos de cada espécie — e cada uma delas tem um tipo, uma referência, um modelo.
Segundo o raciocínio aristotélico (e usando um exemplo já empregado por Sober), esse princípio apresenta dois problemas bem notáveis:
1. Uma onça-pintada tem um filhote negro. O tipo da espécie Panthera onca é amarelado com manchas negras. Portanto, o filhote não possui a essência da mãe, e será uma espécie diferente. A mãe estará amamentando outra espécie.
2. De repente se descobre uma onça vivendo em Marte. Ela jamais viu a Terra, e nenhuma onça terrestre jamais viu nem conheceu essa criatura. Entretanto, são da mesma espécie, pois têm a mesma essência.

Esses exemplos são bem simplistas, e carecem de muitos desdobramentos filosóficos, mas servem para ilustrar, sem muita perda, a deficiência dessa escola de classificação. Isso, claro, não tira seu mérito na história natural: o próprio Carolus Linnaeus a usou quando propôs seu Systema Naturae, em 1758. O que ele fez foi propor um conjunto de regras e categorias para classificação, que foi extremamente útil numa época de explosão de pesquisas e publicações sobre a biodiversidade mundial.
- A Escola Evolutiva

Coincidências à parte, cem anos após a obra-prima de Linnaeus, Charles Darwin e Alfred Wallace chegavam a conclusões muito parecidas, e Darwin, com seu On the Origin of Species (“A Origem das Espécies”), lançou uma das pedras fundamentais da Ciência contemporânea em 1859.
{kind=link}
Não imediatamente, mas pouco a pouco (pois sua teoria, inicialmente tida como anticristã, chegou a ser satirizada), suas ideias passaram a ser consideradas mais seriamente, e já no início do século XX, com o neodarwinismo, tinha se tornado parte do referencial teórico da Biologia moderna, ciência que começava a adquirir autonomia.
{kind=link}
Ainda nessa época, os trabalhos de Wegener (principalmente Die Entstehung der Kontinente und Ozeane, “A Origem dos Continentes e dos Oceanos”) propulsionaram as pesquisas em Biogeografia e Paleontologia: muitos tentavam, então, observar a Terra com um olhar histórico, tentando reconhecer padrões e entender a biodiversidade atual como um desenrolar temporal.
{kind=link}

O impacto que isso trouxe à sistemática e à sociedade em geral foi enorme (a própria Igreja admitiu oficialmente a evolução em 1996). Assim, alguns eminentes taxonomistas e sistematas das décadas de 20, 30 e 40 (entre os quais Mayr e Huxley) começaram a imaginar que as classificações deveriam refletir a evolução de alguma forma. Assim surgia a escola evolutiva (Figura 7.2a).

Uma das controvérsias que surgiram dessa escola estava no fato de que a definição dos grupos taxonômicos dependia dos “grados”, passos evolutivos considerados importantes. Mas... Como saber o que é “importante”?
- A Escola Fenética
Na década de 60, a insatisfação com a escola evolutiva culminou com o surgimento de outra, proposta por Sokal e Sneath. Nessa época, os computadores passavam a ser acessíveis, e os cálculos poderiam ser acelerados consideravelmente. Os feneticistas propunham que, quanto mais parecidas fossem as espécies, mais próximas elas deveriam estar em um cladograma. Portanto, se fossem codificadas em códigos binários todas suas características morfológicas (e depois dos anos 80, moleculares também), seria possível a montagem de grupos inclusivos de similaridade (Figura 7.2b).
A grande vantagem trazida pelos feneticistas foi a robustez de um método, bem menos subjetivo que os empregados na escola evolutiva.
No entanto, apresenta um problema semelhante ao da escola essencialista: será que a mera semelhança pode nos dizer se uma determinada espécie é parente próximo de outra? Muitos feneticistas sabiam dessas limitações, mas não se importavam: para eles, o importante era organizar as espécies de forma consistente. Dado o contexto científico-cultural presente na sociedade acadêmica, sobretudo, esse método passou a sofrer rejeição; afinal, se não se podia agrupar espécies numa tentativa de descobrir seus parentescos, muitos dos potenciais usos da sistemática iriam ralo abaixo.
- A Escola Cladística
 Ironicamente, mais de uma década antes da insatisfação dos sistematas com as escolas existentes, o grande entomólogo germânico Willi Hennig já havia publicado seu Grundzüge einer Theorie der phylogenetischen Systematik, onde aperfeiçoava um dos métodos mais simples e geniais já criados para a sistemática, que lançava raízes no início do século XX. Entretanto, essa técnica em alemão ficou oculta até 1966, quando foi publicada uma versão traduzida para o inglês: daí em diante, o método ganhou adeptos ferrenhos e se disseminou rapidamente. Ainda viria a sofrer muitas críticas e reformas, mas o conceito central permanece há quase 40 anos (Figura 7.2c).
Ironicamente, mais de uma década antes da insatisfação dos sistematas com as escolas existentes, o grande entomólogo germânico Willi Hennig já havia publicado seu Grundzüge einer Theorie der phylogenetischen Systematik, onde aperfeiçoava um dos métodos mais simples e geniais já criados para a sistemática, que lançava raízes no início do século XX. Entretanto, essa técnica em alemão ficou oculta até 1966, quando foi publicada uma versão traduzida para o inglês: daí em diante, o método ganhou adeptos ferrenhos e se disseminou rapidamente. Ainda viria a sofrer muitas críticas e reformas, mas o conceito central permanece há quase 40 anos (Figura 7.2c).
De forma simplificada, a sistemática filogenética procura agrupar organismos por novidades evolutivas compartilhadas (sinapomorfias), sendo que as semelhanças presentes há mais tempo na linhagem não devem ser consideradas (simplesiomorfias); afinal, não podemos agrupar uma minhoca com um marisco só porque são invertebrados: não possuem coluna vertebral, é verdade... Mas também não a possuem os pepinos-do-mar, os ácaros, os líquens, os vírus e as cadeiras de plástico — o que não quer dizer que eles tenham um ancestral comum que seja exclusivo deles.
Que tal mais um exemplo? Vamos a um bastante polêmico, abordado de forma superficial. Considere, portanto, que tem às mãos um conjunto de cinco animais, como mostra a Figura 7.1:

Figura 7.1 Um esturjão (E), uma corruíra (C), um jacaré-de-papo-amarelo (J), um teiú (T) e uma paca (P); o esturjão foi escolhido para termos um critério de comparação (chamado de grupo-externo). Se os agrupássemos apenas com base na semelhança (feneticamente), teríamos um grupo T+J. Mas não podemos fazer isso: as semelhanças entre T e J (escamas, ovos, quatro pernas etc.) não são exclusivas deles; por que, então, deveriam formar um conjunto? Tudo que eles têm de igual, C também tem, embora tenha sofrido ainda mais mudanças. Entretanto, as características que P compartilha com C (desconsiderando a endotermia, para simplificar o exemplo), também compartilha com T e J.
Fonte: Thinkstock / CEPA
No fim das contas, ainda temos um sistema mais simples. Observe:
| Grupos | |
| Fenética |
Sistemática Filogenética |
| T+J+C+P | T+J+C+P |
| T+J | C |
| C | P |
| P | |
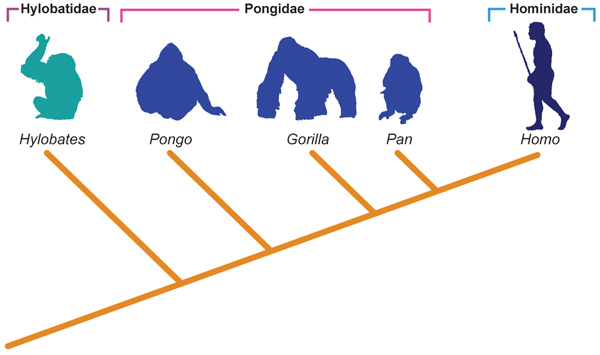
Figura. 7.2a Sistemática e a filogenia dos Hominoidea - Escola Evolutiva.
Fonte: Adaptado de Hickman et al., 2006.
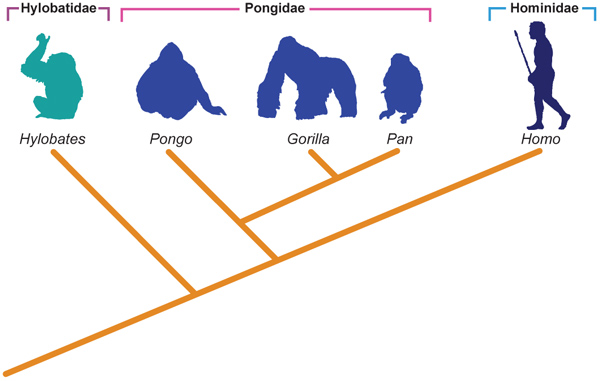
Figura 7.2b Sistemática e a filogenia dos Hominoidea - Escola Fenética.
Fonte: Adaptado de Hickman et al., 2006.
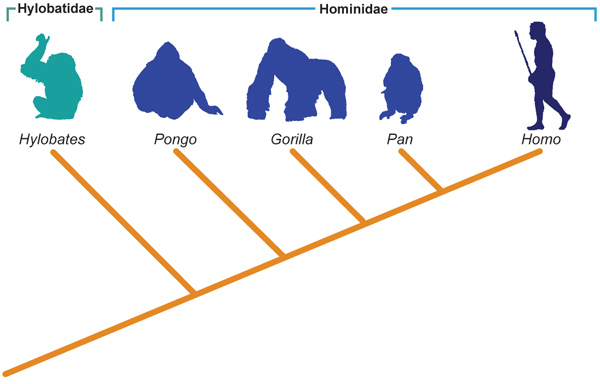
Figura 7.2c Sistemática e a filogenia dos Hominoidea - sistemática Filogenética.
Fonte: Adaptado de Hickman et al., 2006.
- A Evolução na Sistemática Filogenética
Quando se olha para um cladograma, ou árvore filogenética, é necessário entender a simbologia. O tempo transcorre verticalmente, sendo que o topo é mais recente e a base, mais antiga. Horizontalmente temos a diversidade de grupos ao longo do tempo; a menos que haja extinções, casos nos quais a linha se interrompe antes do topo (como o táxon E, na Figura 7.3). Perceba, na figura, que cada nó é o encontro de pelo menos dois ramos (linhas), e representa o ancestral comum e exclusivo de um grupo monofilético, que encerra todos os descendentes daquele ancestral. Por isso, olhando a Figura 7.3, podemos dizer que o grupo BCD é monofilético, mas não os grupos CD, EFG ou ABG, por exemplo.
Talvez fique mais claro se organizarmos esses organismos em um diagrama de Euler-Venn, mais familiar, geralmente, aos alunos (Figura 7.4). Perceba que os conceitos são praticamente os mesmos (repare na inclusividade!), só sendo alterada a maneira de representação.
A sistemática atual não trabalha com táxons (gêneros, famílias etc.) que não sejam monofiléticos. Quando se percebe que um grupo tradicional e antigo não deve ser monofilético após uma série de estudos e análises filogenéticas, os sistematas são obrigados a escolher nomes novos para os grupos estudados.

Figura 7.3 Esquema representando um cladograma. Grupos monofiléticos: ABCDEFG, BCDEFG, BCD, BC, FG, A, B, C, D, E, F, G. O tempo transcorre na vertical; portanto, o ancestral de FG é mais recente que o de BCDEFG, que, por sua vez, pode ser considerado um dos ancestrais de FG.
Fonte: CEPA

Figura 7.4 Diagrama de Euler-Venn representando a mesma árvore vista na Figura 7.3.
Fonte: CEPA
2. Mãos à Obra
 Treinamento: assista à aula interativa da professora Sônia Lopes.
Treinamento: assista à aula interativa da professora Sônia Lopes.
Aperfeiçoamento: quando ocorrer um caso no qual algumas observações contam a favor e outras contra, vence a hipótese com menor conflito. Veja o exemplo abaixo:
| Táxon | Observação | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| X* | 0 | 0 | 0 | 0 | 0 | 0 |
| A | 0 | 0 | 0 | 0 | 1 | 0 |
| B | 0 | 0 | 1 | 1 | 0 | 1 |
| C | 0 | 1 | 1 | 1 | 0 | 1 |
| D | 0 | 1 | 0 | 1 | 0 | 0 |
| Grupos | (ABCD) | (CD) | (BC) | (BCD) | (A) | (BC) |
*Grupo Externo.
Nesse exemplo, temos uma “votação”:
| A: compatível com todos; | ABCD: compatível com todos; | |
| B: compatível com todos; | BCD: compatível com todos; | |
| C: compatível com todos; | BC: incompatível com CD; | |
| D: compatível com todos; | CD: incompatível com BC. |
No caso, como BC tem dois “votos” contra um de CD, teremos um cladograma assim:

Atividade 1 - Cladograma
Envio de Arquivo: observe os insetos abaixo (Netaus niger é o grupo-externo): todos são fêmeas adultas. Esta atividade tem três etapas, que devem constar no seu arquivo:
1. Monte uma matriz de características observáveis, com pelo menos 5 colunas:
- Coloque a legenda das colunas conforme o exemplo:
Largura dos braços:
a. Braços largos, com cerca de 1 m de largura;
b. Braços estreitos, com cerca de 20 cm de largura.
2. Faça um cladograma, colocando as sinapomorfias (como nos exemplos vistos em “Mãos à Obra”, acima). No lugar dos terminais, você deve usar o nome dos besouros.
3. Você alteraria o nome do gênero de algum deles? Se sim, tente explicar com base no cladograma.

3. Finalizando
 Você já deve ter percebido a importância dos estudos sobre a classificação (e não só dos animais!). Extrapolando um cladograma, fazendo inferências filogenéticas, podemos “retrover” acontecimentos que devem ter ocorrido há milhões de anos; mas, partindo de supostas relações de parentesco, podemos também “prever” algumas características de determinados táxons. Quando você olha para um cladograma e localiza o ancestral comum mais recente entre as vespas-caboclas e as abelhas-de-mel, pode arriscar que um descendente qualquer desse ancestral, escolhido ao acaso, deve ter também ferrão como as abelhas-de-mel e as vespas-caboclas.
Você já deve ter percebido a importância dos estudos sobre a classificação (e não só dos animais!). Extrapolando um cladograma, fazendo inferências filogenéticas, podemos “retrover” acontecimentos que devem ter ocorrido há milhões de anos; mas, partindo de supostas relações de parentesco, podemos também “prever” algumas características de determinados táxons. Quando você olha para um cladograma e localiza o ancestral comum mais recente entre as vespas-caboclas e as abelhas-de-mel, pode arriscar que um descendente qualquer desse ancestral, escolhido ao acaso, deve ter também ferrão como as abelhas-de-mel e as vespas-caboclas.
{kind=link}
{kind=link}
A sistemática filogenética é a escola mais bem aceita, atualmente, na comunidade científica, mas não é unanimidade: como você deve ter percebido, dificilmente nossas observações sobre as características de um grupo de espécies são todas coincidentes; quando há conflito, optamos pela maioria das ditas “evidências” (chamadas observações ou caracteres): esse é, de forma rasa, o princípio da parcimônia, corriqueiro na prática científica. Quando uma característica se mostra presente em grupos não monofiléticos não pode ser considerada uma sinapomorfia, uma homologia: trata-se, então, de uma homoplasia (que pode ser uma convergência ou uma reversão a uma condição ancestral).
Mas nada nos garante que a evolução seja parcimoniosa, que ela opte pelos caminhos mais curtos e diretos; e esse é um dos principais ataques contra a sistemática filogenética, usados pelos proponentes de outras escolas (como a “probabilística”), que insistem que nem sempre o mais óbvio é o mais provável. Essa discussão é inflamada no meio acadêmico, e deve haver mais de um exemplo no qual determinadas pessoas, que em outras ocasiões seriam bons colegas, acabam se vendo como inimigos ferrenhos.
Para obtenção desses cladogramas podem ser usadas as mais variáveis fontes de dados: morfologia, fisiologia, bioquímica, citologia, genética, biologia molecular, histologia, comportamento... Certamente “não vale qualquer coisa”, mas muita coisa vale!
Como última nota — e que sirva de consolo para você que suou na montagem do cladograma desta semana! —, a resolução dessas matrizes de táxons e observações torna-se rapidamente algo insolúvel para uma mente humana, o que faz com que supercomputadores e programas eficientes se tornem imprescindíveis (e mesmo assim, para encontrar resultados aproximados).
Só a título de comparação: com quatro táxons, temos um número de árvores possíveis igual a 15. Caso fossem cerca de 70 táxons, o número seria de míseros... (tente falá-lo)... 10120! Tente imaginar, então, o número de cladogramas possíveis para todas as quase dois milhões de espécies conhecidas... Bem, eu desisto, vou dormir. Se você por acaso chegar à resposta correta, avise à NASA para que jogue todos seus computadores no lixo e o contrate!
4. Ampliando os Conhecimentos
 Amorim, D.S., 1994. Elementos Básicos de Sistemática Filogenética. Sociedade Brasileira de Entomologia.
Amorim, D.S., 1994. Elementos Básicos de Sistemática Filogenética. Sociedade Brasileira de Entomologia.
Goloboff, P.A., J S. Farris, K.C. Nixon, 2008. TNT, a free program for phylogenetic analysis. Cladistics 24(5): 774-786. [talvez o melhor programa para construção de árvores filogenéticas já criado. Pode ser baixado em: http://www.zmuc.dk/public/phylogeny/TNT/. É gratuito, mas o preço é que está em inglês!]
Tree of Life, 2007. Tree of Life Web Project. http://tolweb.org/tree/, acesso em 02/2011. [se você ainda está calculando o número de árvores possíveis para toda a vida, pare. Os caras já estão na sua frente! Esse site propõe uma grande aproximação para classificar tudo que vive ou já viveu na Terra.]
University of California Museum of Paleontology. What Did T. rex Taste Like? http://www.ucmp.berkeley.edu/education/explorations/tours/Trex/phych1A.html, acesso em 02/2011. [este é para quem adorou a atividade da professora Sônia Lopes! A desvantagem é estar em inglês.]
5. Sugestão de Atividades
 As aulas interativas que você mesmo já espiou são instrumentos muito didáticos e lúdicos, e teriam grande apelo se aplicados junto aos alunos.
As aulas interativas que você mesmo já espiou são instrumentos muito didáticos e lúdicos, e teriam grande apelo se aplicados junto aos alunos.
Nada impede, porém, que você mesmo use figuras simples (como monstrinhos ou animais caricatos simples, com poucos traços) para treinamento com seus alunos, numa proposta similar à que você fez na seção “Mãos à Obra” deste tópico.
Atividade 2 – Fórum de Discussão sobre Taxonomia
“A taxonomia deve ser considerada uma ciência? Por quê?” Além de colocar sua resposta, não se esqueça de ponderar sobre a de um colega e colocar sua opinião a respeito.
6. Bibliografia
 Amorim, D.S. Fundamentos de Sistemática Filogenética, 2002. Editora Holos.
Amorim, D.S. Fundamentos de Sistemática Filogenética, 2002. Editora Holos.
Pough, F.H., C.M. Janis, & J.B. Heiser, 2003. A Vida dos Vertebrados. Editora Atheneu.
Ruppert, E., Fox, R.S. & Barnes, R.D., 2005. Zoologia dos Invertebrados - Uma abordagem funcional-evolutiva. Editora Roca.
Hickman Jr., C.P., L.S. Roberts, A. Larson, 2004. Princípios Integrados de Zoologia. Editora Guanabara-Koogan.
7. Anexos
 A seguir, sites para aulas interativas e um texto sobre sistemática filogenética. Podem ser utilizados em sala de aula e/ou para o estudo do professor com o fim de revisar e aprofundar seus conhecimentos e de seus alunos.
A seguir, sites para aulas interativas e um texto sobre sistemática filogenética. Podem ser utilizados em sala de aula e/ou para o estudo do professor com o fim de revisar e aprofundar seus conhecimentos e de seus alunos.
http://www.ib.usp.br/md/arquivos/evolucao.FLA.swf
http://www.johnkyrk.com/evolution.pt.html
http://www.ib.usp.br/md/arquivos/evolucao.FLA.swf
http://www.ib.usp.br/microgene/atividades0popup.php?Arquivo=atividades-2-Arquivo.swf
http://www.ib.usp.br/microgene/atividades0popup.php?Arquivo=atividades-3-Arquivo.swf
PDF - Termos e Conceitos sobre Sistematica Filogenetica - Prof. Yuri Leite – UFES.