Texto 8 – Ciclo Celular e Mitose
| Sitio: | Moodle USP: e-Disciplinas |
| Curso: | 01 - Biologia Celular |
| Libro: | Texto 8 – Ciclo Celular e Mitose |
| Imprimido por: | Usuário visitante |
| Día: | miércoles, 17 de julio de 2024, 07:56 |
Descripción
Texto 8 – Ciclo Celular e Mitose
Tabla de contenidos
- 1. Iniciando a conversa
- 2. Introdução
- 3. O ciclo celular
- 4. A mitose
- 5. A periodicidade do ciclo celular
- 6. A prófase
- 7. As alterações no citoesqueleto e na compactação da cromatina durante a mitose
- 8. A prometáfase e a metáfase
- 9. Morfologia cromossômica
- 10. Os telômeros
- 11. A anáfase
- 12. A telófase e a citocinese
- 13. Divisão celular na célula vegetal
- 14. Falha do controle do ciclo celular e o desenvolvimento de tumores
- 15. Câncer
- 16. Bibliografia
- 17. Mãos à obra
- 18. Ampliando os conhecimentos
1. Iniciando a conversa
Os objetivos específicos do Segundo Tópico do Tema 4 são:
-
Entender as fases do ciclo celular;
-
Compreender os processos envolvidos na mitose e reconhecer suas fases;
-
Reconhecer as possíveis falhas do controle do ciclo celular e o desenvolvimento de tumores, incluindo os cancerígenos.
No Ensino Médio, trazer aspectos do cotidiano para as discussões na escola sempre é uma boa estratégia para motivar os estudantes. No caso da temática do ciclo celular, a ocorrência de algumas doenças pode ser um desses aspectos do dia a dia.
O ciclo celular controla como e quando ela deve se desenvolver e proliferar em harmonia com o restante do organismo. Falhas no controle deste ciclo favorecem a formação de células que se desenvolvem de modo desenfreado, sem levar em conta o “bem estar” das células ao seu redor, os tumores.
Assista à animação que apresenta a formação de um tumor e a ocorrência de metástase: .
Podemos ver nessa animação como as células tumorais se proliferam. Essa proliferação descontrolada está relacionada a alterações do ciclo celular, como dito anteriormente. O ciclo celular e a mitose são justamente os temas do presente tópico.
Também iremos abordar em maior detalhe uma das doenças mais presentes na população humana: o câncer.
2. Introdução
“Omnis cellula e cellula” foi um importante conceito estabelecido por Rudolf Virchow em 1858, que estabelece o princípio de que todas as células se originam de outras pré-existentes, complementando a teoria celular proposta anos antes. Para tanto, uma sequência de eventos é desencadeada na célula, que envolvem tanto modificações morfológicas quanto fisiológicas e culminam no processo de divisão celular.
Este é o principal mecanismo de proliferação celular, possuindo duas funções: a produção de novas células de igual conteúdo gênico e citoplasmático, sendo esta a divisão mitótica, e a produção de gametas e variabilidade genética, resultado de uma outra modalidade de divisão celular que é a divisão meiótica, que será abordada em outro curso.
3. O ciclo celular
Para organismos unicelulares e células de linhagem somática dos metazoários, o ciclo celular mitótico pode ser compreendido como uma sucessão de etapas controladas por pontos de checagem, que irão garantir a ocorrência correta do processo, impedindo que a célula se divida em um momento inapropriado ou passe imperfeições às células filhas. A peridiocidade deste ciclo é determinada pela ativação e desativação cíclica de proteínas-chaves presentes no citosol, das quais falaremos mais adiante. Este processo é normalmente dividido temporalmente em quatro grandes etapas sequenciais (Fig. 1): a fase G1, a fase S e a fase G2, que compõem a intérfase, seguidas da fase M, que corresponde à mitose propriamente dita.
Existem, ainda, linhagens proliferativas de células cuja ocorrência da divisão é menos frequente, permanecendo longos períodos fora do ciclo celular. Dizemos, então, que estas células estão na fase G0. Elas podem permanecer nesta fase, em um estado diferenciado onde não mais se dividem, ou, de acordo com a necessidade do organismo, podem sair de G0 e retornarem à fase G1.
Neste período G1 da intérfase, a célula apresenta elevados níveis de transcrição e síntese protéica, bem como aumento na taxa de crescimento celular. Ao final desta fase, há o primeiro ponto de checagem para a passagem para a fase S, onde é verificado se a célula atingiu o tamanho adequado para a continuidade do processo e se o ambiente está favorável para a divisão. Após esta verificação a célula segue o ciclo, dando início à duplicação de todo o material genético. Uma vez assumido o compromisso pela célula para a continuidade do ciclo após o ponto de checagem, mesmo a remoção dos sinais para crescimento e divisão não levam a parada do ciclo. A fase S é caracterizada pela duplicação de todo o DNA presente no núcleo, preparando-o para a divisão celular.
Seguindo a duplicação do DNA adentramos a fase G2, bastante semelhante à fase G1, diferindo apenas na checagem realizada em seu final que, neste caso, envolve a verificação do término da duplicação e da integridade do material genético, garantindo à célula tempo suficiente para a ativação da maquinaria de reparo do DNA antes de prosseguir o ciclo celular.

Figura 1: Fases principais do ciclo celular mostrando os pontos de checagem (A) e um exemplo de duração de cada uma delas (B). Note que a célula poderá sair do ciclo definitiva ou temporariamente entrando na chamada fase G0.
Fonte: POLLARD, T.D. & EARNSHAW, W.C. – Biologia celular. Rio de Janeiro, Elsevier, 2006.
4. A mitose
A fase M (de Mitose) ocorre em um curto período de tempo, quando comparada à intérfase. Entretanto, as modificações decorrentes desta etapa são mais intensas, levando a uma reestruturação quase completa da organização celular.
A mitose é classicamente dividida em cinco fases distintas: prófase, prometáfase, metáfase, anáfase e telófase. Vale lembrar que apesar da divisão em fases, todos os processos descritos são contínuos e que a mitose é sempre equitativa, ou seja, mantém, em suas células-filhas a mesma quantidade de DNA das células-mães.
5. A periodicidade do ciclo celular
A periodicidade do ciclo celular está associada, como mencionado anteriormente, a proteínas-chaves presentes no citossol, mais especificamente a uma família de proteínas denominadas Cdks (do inglês ciclin dependent kinase ou quinase dependente de ciclina), cuja atividade cíclica ao longo do ciclo celular fará com que a célula progrida no ciclo. Tal atividade é resultado da interação das Cdks com conjuntos de complexos de enzimas e outras proteínas, dentre as quais a proteína ciclina é conhecida como a mais importante reguladora das Cdks.
Como o próprio nome diz, as Cdks necessitam de uma associação íntima com as ciclinas para que sua atividade como quinase (fosforilação) seja possível. Os níveis de Cdks se mantêm constantes durante todo o ciclo celular. Entretanto, as ciclinas sofrem um processo de síntese e degradação ao longo do ciclo, tornando periódica a ativação do complexo Cdk-ciclina.
A periodicidade na ativação deste complexo faz com que sejam desencadeados inúmeros eventos do ciclo celular. Cada um das etapas do ciclo celular é regulada por um complexo Cdk-ciclina específico, sendo o complexo G1-Cdk presente ao longo da fase G1; G1/S-Cdk, responsável pelo ponto de verificação para passagem a fase S; S-Cdk, necessária para início da duplicação do DNA; M-Cdk, responsável por promover inúmeros eventos durante a mitose, como condensação cromossômica, desorganização do envoltório nuclear, remodelação do citoesqueleto, reorganização do complexo de Golgi e do retículo endoplasmático.
Cada um desses complexos apresenta, no caso de vertebrados, pelo menos uma ciclina e uma Cdk exclusiva para cada etapa, demonstrando a elevada complexidade da regulação do ciclo celular. Acredita-se que a ciclina não tem por função simplesmente ativar as Cdks, mas também direcionar sua ação, levando à fosforilação de substratos específicos, como vimos no caso da M-Cdk que desencadeiam várias respostas fisiológicas durante a fase M (Fig. 2).
 envolvidos na regulação das fases do ciclo celular")
Figura 2: Principais complexos protéicos (ciclinas/Cdk) envolvidos na regulação das fases do ciclo celular.
Fonte: KARP, G. - Cell molecular biology. New York, J. Wiley, 1996.
6. A prófase
Como já mencionado, a mitose tem início na prófase (Fig. 3). Na prófase ocorrem os primeiros eventos da divisão, tais como o início da migração dos centrossomos (que se dividiram na fase S) para pólos opostos; início da desestruturação do envoltório nuclear e da condensação dos cromossomos.

Figura 3: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em prófase vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006. ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
Os centrossomos (cada um constituído por um par de centríolos e por várias proteínas associadas) funcionam como centros organizadores dos microtúbulos que constituem o aparelho mitótico. O aparelho mitótico é constituído, basicamente, pelo fuso mitótico, formado por fibras polares (que terminam no equador da célula em divisão) e por fibras cromossômicas ou cinetocóricas (que se ligam aos cromossomos).
Além do fuso, o aparelho mitótico apresenta os chamados ásteres, constituídos por microtúbulos curtos que irradiam dos centros celulares (Fig. 4).

Figura 4: Desenho esquemático do aparelho mitótico, formado pelo fuso mitótico, os ásteres e os entros celulares (centríolos e material pericentriolar com proteínas associadas).
Fonte: KARP, G. - Cell molecular biology. New York, J. Wiley, 1996.
7. As alterações no citoesqueleto e na compactação da cromatina durante a mitose
Vale lembrar que, quando a célula entra em divisão, os cromossomos já estão compactados no nível de alças (terceiro nível de compactação) da intérfase e iniciam mais um nível de compactação no início da mitose (Fig. 5).
O citoesqueleto microtubular interfásico da célula começa então a se reorganizar para formar o aparelho mitótico. Portanto, o aparelho mitótico nada mais é do que a rede de microtúbulos da célula interfásica reestruturados. Durante este período da prófase, ainda há atividade gênica e produção de proteínas e, portanto, o nucléolo ainda pode ser visualizado.

Figura 5: Esquema geral dos diferentes níveis de compactação da cromatina com destaque para o nível final de compactação existente no cromossomo metafásico (E-F).
Fonte: LODISH, H, BERK, A., MATSUDAIRA, P., KAISER C.A., KRIEGER M., SCOTT M.P., ZIPURSKY, S.L. & DARNELL, J. Biologia Celular e Molecular. 5a edição. Porto Alegre, Artmed, 2005.
8. A prometáfase e a metáfase
Na prometáfase (Fig. 6) os centrossomos continuam a migração para os pólos opostos da célula e alguns microtúbulos do fuso encontram-se ligados a regiões específicas dos cromossomos denominadas de cinetócoros (fibras cinetocóricas), localizadas nos centrômeros (Fig. 4), que correspondem a uma pequena constrição em cada cromossomo duplicado, sendo cada uma das cópias denominada de cromátide. Segue-se a desestruturação do envoltório nuclear, bem como a condensação dos cromossomos, que estão mais próximos do plano equatorial da célula. A partir deste ponto, a atividade gênica é extremamente reduzida e o nucléolo desaparece.

Figura 6: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em prometáfase vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006. ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
Na metáfase os centrossomos estão localizados nos pólos opostos da célula e todos os cinetócoros estão ligados aos fusos (Fig.4, 6); o envoltório nuclear está completamente desestruturado e os cromossomos, já se encontram no plano equatorial da célula. Nesta fase, o DNA cromossômico atinge seu nível máximo de compactação, formando a fibrila de 700 nm (quarto nível de compactação), de modo que as duas cromátides unidas têm um diâmetro total de 1400 nm.

Figura 7: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em metáfase vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006. ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
Esta fibrila é resultado do enrolamento do arcabouço protéico, formando um helicóide com as alças de cromatina espalhadas nos diferentes planos. Este nível de compactação permite a visualização mais evidente dos cromossomos onde se pode identificar duas regiões bem características em cada um deles: um centrômero e dois telômeros (regiões terminais dos cromossomos)(Fig. 7).

Figura 8: Cromossomos metafásicos evidenciando os centrômeros e os telômeros.
Fonte: Composição de: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008 e Site: http://www.cakens.com/research/understanding-the-mechanisms-of-ageing-worth-a-nobel-prize/ acesso em 10/09/2010.
A região centromérica, acima citada, é composta basicamente por seqüências repetitivas de DNA, as quais permanecem associadas a um complexo protéico, também já mencionado, denominado cinetócoro, o local de associação dos fusos durante a divisão celular. Devido a essa associação, a região centromérica é de extrema importância para a célula, uma vez que a ligação correta dos fusos aos cinetócoros garantirá o sucesso da divisão celular, permitindo a perpetuação das diferentes linhagens de células.
9. Morfologia cromossômica
Como já relatado, na metáfase é possível uma boa visualização dos cromossomos, devido à sua máxima condensação. Assim, verifica-se que cada cromossomo é formado, na verdade, por dois cromossomos idênticos, as cromátides (resultado da duplicação na fase S), ainda unidos pelo centrômero.
A posição do centrômero nos permite classificar morfologicamente os cromossomos metafásicos em quatro tipos distintos (Fig. 9):
-
metacêntricos: centrômero aproximadamente centralizado, formando dois pares de braços cromossômicos (partes do cromossomo separadas pelo centrômero) de igual tamanho;
-
submetacêntricos: centrômero levemente deslocado da região mediana, formando braços pouco mais curto que os outros;
-
acrocêntricos: centrômero muito próximo a uma das extremidades, formando dois braços extremamente curtos e os outros bastante longos;
-
telocêntricos: centrômero localizado na extremidade do cromossomo, formando um par de braços longos.

Figura 9: Tipos morfologicos dos cromossomos metafásicos: metacêntrico (a), submetacêntrico (b), acrocêntrico (c) e telocêntrico (d).
Fonte: Original
O fato de os cromossomos metafásicos estarem mais visíveis ao microscópio, como mencionado acima, permite não apenas identificar os tipos de cromossomos presentes, como também o seu número e morfologia. Através da análise fotográfica de várias metáfases ao microscópio, é possível montar um arranjo perfeito dos vários pares de cromossomos presentes, numa disposição que se denomina cariótipo (Fig. 10).
O estudo do cariótipo humano tem uma enorme importância clínica permitindo a detecção eventual de aberrações cromossômicas, envolvendo alterações no número e/ou morfologia dos cromossomos, responsáveis por uma infinidade de síndromes genéticas existentes no homem. A identificação de problemas no cariótipo é fundamental, por exemplo, nos vários procedimentos experimentais que se utiliza no aconselhamento genético de casais que desejam saber, por exemplo, qual seria o risco de terem filhos com problemas genéticos de natureza cromossômica.
")
Figura 10: Imagem de uma metáfase de uma célula humana vista ao microscópio (A). Montagem dos mesmos cromossomos na forma de cariótipo (B). A técnica de preparação utilizada resulta na formação de um padrão de bandas específico para cada par cromossômico, facilitando sua identificação.
Fonte: Composição de: site: http://biochem.co/2008/07/dna-introduction-to-deoxyribonucleic-acid/ acesso em 10/09/2010 e site: http://www.medscape.com/viewarticle/505220_7 acesso em 10/09/2010
10. Os telômeros
Os telômeros (Fig. 8), por sua vez, compõem as regiões terminais de todos os cromossomos e muitas são as hipóteses sobre qual a real função dessas regiões. De modo geral, sabe-se que tais regiões irão garantir a perpetuação dos cromossomos na divisão celular e são, assim como os centrômeros, compostos por seqüências repetitivas de DNA. Entretanto, diferentemente das regiões centroméricas, estas seqüências são altamente conservadas nas diferentes espécies, o que denota uma grande importância associada a esse segmento.
Ainda é assunto de discussão entre os pesquisadores se as regiões teloméricas atuam como regiões protetoras das extremidades dos cromossomos e qual é a contribuição efetiva dessas regiões no processo de envelhecimento celular, ao qual parecem estar intimamente ligadas.
11. A anáfase
Ao final da metáfase ocorre o terceiro ponto de checagem, no qual é verificado se todos os cinetócoros estão perfeitamente ligados ao fuso (Fig. 1). Em caso positivo, a célula segue para a anáfase com a ativação do APC (complexo promotor de anáfase ou, do inglês, anaphase promoting complex), o qual irá atuar na coesina, proteína que mantém as cromátides irmãs unidas, inativando-a (Fig. 11). Inicia-se então a anáfase propriamente dita (Fig. 12) com a separação das cromátides irmãs por dois processos distintos, porém simultâneos, denominados anáfases A e B (Fig. 13).
A anáfase A tem por característica o encurtamento dos fusos ligados aos cinetócoros, levando, assim, à separação das cromátides irmãs. A anáfase B, por sua vez, tem a participação dos fusos polares, que são fusos oriundos dos pólos opostos, mas que não estão ligados a nenhum cinetócoro.
Inicia-se então a anáfase propriamente dita com a separação das cromátides irmãs por dois processos distintos, porém simultâneos, denominados anáfases A e B (Fig. 10).
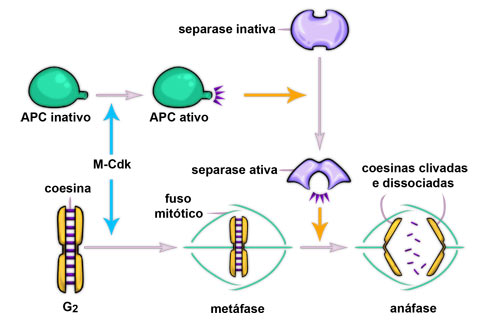
Figura 11: Ação do complexo promotor de anáfase (APC) na inativação da coesina, permitindo a separação das cromátides irmãs na anáfase.
Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.

Figura 12: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em anáfase vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006. e ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.

Figura 13: Os dois tipos de movimentos que ocorrem durante a anáfase levando à separação das cromátides irmãs (anáfase A e anáfase B).
Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
As extremidades livres destes fusos se sobrepõem na região da placa equatorial da célula. O deslizamento dos fusos polares em sentidos opostos acarreta num distanciamento dos centrossomos, auxiliando a separação das cromátides irmãs. Acredita-se que esse distanciamento seja mediado por proteínas motoras presentes na região da sobreposição dos fusos polares. Sendo assim, a anáfase é constituída de dois movimentos simultâneos, um de encurtamento dos fusos e outro de distanciamento dos centros celulares que otimizam o processo de separação das cromátides irmãs.
12. A telófase e a citocinese
A telófase (Fig. 14) corresponde ao processo inverso que ocorre na prófase. Ocorre a reestruturação do envoltório nuclear, a descondensação dos cromossomos que retornam ao nível de alças, o reaparecimento do nucléolo e a retomada da atividade gênica. Além destes processos, há a formação de um anel contrátil de microfilamentos associados à miosina no plano equatorial da célula, dando início à citocinese, processo de divisão do citoplasma, que irá finalizar o processo mitótico (Fig. 15, 16).

Figura 14: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em telófase vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006 e ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.

Figura 15: Esquema geral das várias fases da mitose. A imagem central mostra uma célula em citocinese vista ao microscópio de fluorescência (cromossomos em vermelho, fuso de divisão com os microtúbulos em verde).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006 e ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.

Figura 16: Esquema de uma citocinese mostrando o anel contráctil formado por feixes circulares de microfilamentos associados à miosina (A). Imagens de microscopia eletrônica exibindo detalhes do sulco de divisão resultante da atuação do anel contráctil.
Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
13. Divisão celular na célula vegetal
As células vegetais possuem algumas particularidades na fase M. Sua divisão é anastral, ou seja, apesar da existência dos centrossomos e sua migração para pólos opostos, não existem centríolos e nem ásteres. Além disso, sua citocinese é centrífuga, ao contrário da centrípeta das células animais, ou seja, ocorre do interior da célula para fora, ao invés do estrangulamento de fora para dentro das células animais.
A formação da nova parede celular (incluindo a membrana plasmática) a partir da lamela média, é o resultado da fusão de inúmeras vesículas, oriundas, em grande parte, do complexo de Golgi, na região central da célula, onde existe um arcabouço de microtúbulos denominado fragmoplasto. Essa lamela, então, se expande para a periferia até alcançar a membrana plasmática, dividindo a célula em duas. Posteriormente, microfibrilas de celulose irão se depositar na matriz da lamela, completando, assim, a formação de uma nova parede celular (Fig. 17).

Figura 17: Divisão mitótica de uma célula vegetal mostrando detalhes da citocinese e formação da nova parede celular.
Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
14. Falha do controle do ciclo celular e o desenvolvimento de tumores
O controle do ciclo celular é bastante rígido, pois qualquer erro na regulação deste processo pode levar à formação de células defeituosas que, por sua vez, têm o potencial de se reproduzir e passar este defeito para suas células-filhas, podendo ter conseqüências desastrosas para o organismo. Entretanto, erros neste controle ocorrem com freqüência e há inúmeros sistemas de reparo que evitam a proliferação dos mesmos.
Em casos extremos, nos quais este sistema de reparo não é suficiente, a célula pode ser encaminhada para um processo complexo que resulta na assim chamada morte celular programada.
Há casos em que o próprio sistema de reparo é defeituoso, permitindo a perpetuação destas linhagens de células imperfeitas. Muitas vezes, estas células proliferam de modo desordenado, ou seja, elas se dividem independentemente da demanda do organismo, crescendo como células individuais, ao invés de crescerem como um tecido, respeitando suas limitações espaciais estabelecidas pelas células adjacentes. Como resultado desta proliferação desordenada, forma-se uma única massa de células denominada tumor (Fig. 18). Em geral, o tumor é do tipo benigno e não traz grandes prejuízos ao organismo, podendo ser completamente removido cirurgicamente.
 e maligno (invasivo)")
Figura 18: Esquemas mostrando o epitélio de um órgão tubular com a formação de um tumor benigno (circunscrito) e maligno (invasivo).
Fonte: ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
15. Câncer
O câncer é uma doença resultante de um tipo de tumor, denominado tumor maligno (Fig. 19), cujas células se desprendem da lâmina basal e têm o potencial de invadir tecidos adjacentes. Em alguns casos, estas células tumorais adquirem a capacidade de se desprender do tumor primário e entrar tanto na circulação sanguínea quanto na linfática, formando, assim, tumores secundários, distantes do local de formação do tumor original, caracterizando, deste modo, a denominada metástase. Quanto mais o tumor se espalha, mais difícil será eliminá-lo.

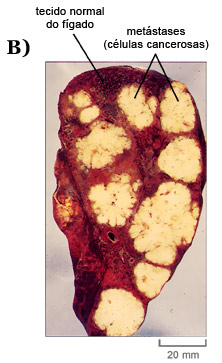
Figura 19: Desenho esquemático da formação de um tumor maligno cujas células se soltam e se espalham por via circulatória podendo se instalar em outras regiões do organismo, num processo denominado metástase (A). Imagem de um fígado humano onde se pode observar células normais e tumorais, decorrentes de metástase (B).
Fonte: Composição de: ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006 e ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
Um dos fatores determinantes para o desenvolvimento do câncer é o ambiental, embora, os hábitos de vida, assim como a predisposição genética do indivíduo, sejam também extremamente relevantes para a ocorrência da doença. Estes fatos explicam a diferença nos tipos de cânceres predominantes em diferentes regiões e populações. Um exemplo típico de uma causa ambiental para o desenvolvimento da doença é o excesso de incidência de radiação ultra-violeta do sol nas células, que pode ocasionar alterações no DNA, levando à formação de um câncer de pele.
De modo geral, a ocorrência do câncer se dá, principalmente, em células da linhagem somática e, portanto, não é hereditária, como no caso do câncer de pele descrito anteriormente. Entretanto, existe a possibilidade desta mutação ocorrer em células da linhagem germinativa, podendo ser passada para a próxima geração. Neste caso, a predisposição para o desenvolvimento da doença existe, mas não necessariamente é suficiente para ocasionar o câncer. Fatores exógenos são, então, indispensáveis para tanto.
Para que as células tumorais se desenvolvam deste modo, as mutações acumuladas devem conferir vantagem adaptativa em relação às demais células do organismo. Diferentes tipos de câncer requerem diferentes combinações de comportamentos chave. Entretanto, podemos listar, de modo geral, alguns destes comportamentos:
-
Dependência reduzida de sinais oriundos de células adjacentes para crescimento, sobrevivência e divisão;
-
Menor propensão de morrerem por morte celular programada;
-
Proliferação sem controle;
-
Células geneticamente instáveis e com alta taxa de mutação;
-
Células anormalmente invasivas;
-
Capacidade de dispersão e proliferação para tecidos distantes (metástase).
No caso do câncer, a melhor opção é sempre a prevenção e não o tratamento. Portanto, um hábito de vida saudável aliado a consultas regulares ao médico assistente podem evitar o surgimento de vários tipos de câncer ou diagnosticá-lo em suas fases iniciais, o que melhora, significativamente, o prognóstico de cura.
16. Bibliografia
ALBERTS, B.; BRAY, O.; HOPKIN, K., JOHNSON A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. Fundamentos da Biologia Celular. 2a. edição. Porto Alegre, Artmed, 2006.
ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. & WALTER, P. - Molecular Biology of the Cell. 5th Edition, New York, Garland, 2008.
KARP, G. - Cell molecular biology. New York, J. Wiley, 1996.
LODISH, H, BERK, A., MATSUDAIRA, P., KAISER C.A., KRIEGER M., SCOTT M.P., ZIPURSKY, S.L. & DARNELL, J. Biologia Celular e Molecular. 5a edição. Porto Alegre, Artmed, 2005.
POLLARD, T.D. & EARNSHAW, W.C. – Biologia celular. Rio de Janeiro, Elsevier, 2006.
site: http://biochem.co/2008/07/dna-introduction-to-deoxyribonucleic-acid/ acesso em 10/09/2010.
Site: http://www.cakens.com/research/understanding-the-mechanisms-of-ageing-worth-a-nobel-prize/ acesso em 10/09/2010.
site: http://www.medscape.com/viewarticle/505220_7 acesso em 10/09/2010.
17. Mãos à obra
Atividade 1 - Não esqueça de responder o questionário.
18. Ampliando os conhecimentos
Assista aos seguintes vídeos:
-
Ciclo celular – visão geral, incluindo G0:
-
Divisão em animação mostrando ligação fuso com cinetócoro e com anáfases A e B:
-
Divisão célula vegetal com formação do fragmoplasto:
-
Mitose em tempo real de Drosophila evidenciando a grande alteração no citoesqueleto: